Abstract
The plant cannabinoids (phytocannabinoids), cannabidiol (CBD), and Δ9-tetrahydrocannabinol (THC) were previously shown to activate transient receptor potential channels of both vanilloid type 1 (TRPV1) and ankyrin type 1 (TRPA1), respectively. Furthermore, the endocannabinoid anandamide is known to activate TRPV1 and was recently found to antagonize the menthol- and icilin-sensitive transient receptor potential channels of melastatin type 8 (TRPM8). In this study, we investigated the effects of six phytocannabinoids [i.e., CBD, THC, CBD acid, THC acid, cannabichromene (CBC), and cannabigerol (CBG)] on TRPA1- and TRPM8-mediated increase in intracellular Ca2+ in either HEK-293 cells overexpressing the two channels or rat dorsal root ganglia (DRG) sensory neurons. All of the compounds tested induced TRPA1-mediated Ca2+ elevation in HEK-293 cells with efficacy comparable with that of mustard oil isothiocyanates (MO), the most potent being CBC (EC50 = 60 nM) and the least potent being CBG and CBD acid (EC50 = 3.4–12.0 μM). CBC also activated MO-sensitive DRG neurons, although with lower potency (EC50 = 34.3 μM). Furthermore, although none of the compounds tested activated TRPM8-mediated Ca2+ elevation in HEK-293 cells, they all, with the exception of CBC, antagonized this response when it was induced by either menthol or icilin. CBD, CBG, THC, and THC acid were equipotent (IC50 = 70–160 nM), whereas CBD acid was the least potent compound (IC50 = 0.9–1.6 μM). CBG inhibited Ca2+ elevation also in icilin-sensitive DRG neurons with potency (IC50 = 4.5 μM) similar to that of anandamide (IC50 = 10 μM). Our findings suggest that phytocannabinoids and cannabis extracts exert some of their pharmacological actions also by interacting with TRPA1 and TRPM8 channels, with potential implications for the treatment of pain and cancer.

Transient receptor potential (TRP) ligand-gated cation channels are expressed in many species ranging from yeast to mammals. TRP channels of vanilloid (TRPV), melastatin (TRPM), and ankyrin (TRPA) types are required for the transduction of sensory stimuli by dorsal root ganglia (DRG) and trigeminal (TG) neurons, some of which mediate noxious thermal or mechanical stimuli, whereas others are polymodal nociceptors. Temperature-sensitive TRP channels are activated by endogenous and exogenous compounds that often excite nociceptors and elicit pain and peripheral inflammation (Levine and Alessandri-Haber, 2007).
TRPV type 1 (TRPV1) is activated by natural products, such as capsaicin and resiniferatoxin, noxious heat temperatures (≥42°C), and low pH (<5.9) (Caterina et al., 1997). A plant cannabinoid (phytocannabinoid), cannabidiol (CBD), also activates and desensitizes TRPV1 (Bisogno et al., 2001; Ligresti et al., 2006), and this effect is responsible for some of the analgesic and anti-inflammatory actions of this compound (Costa et al., 2004, 2007). TRPM8 (Tsavaler et al., 2001) is expressed in both nociceptive and non-nociceptive neurons (Xing et al., 2006). It is gated by low (<25°C) temperatures and chemical cooling compounds such as menthol, eucalyptol, spearmint, WS-3, and icilin (McKemy et al., 2002; Bandell et al., 2004; Chuang et al., 2004), and it is regulated, by being inactivated, in a way opposite to that observed with TRPV1 by protein kinase C-mediated (Premkumar et al., 2005) and protein kinase A-mediated phosphorylation and by the endogenous TRPV1 agonist anandamide (De Petrocellis et al., 2007). TRPA1 (Corey et al., 2004) is found in small-diameter TG and DRG neurons distinct from those expressing TRPM8 but coexpressing TRPV1, substance P, calcitonin gene-related peptide (CGRP), and bradykinin receptors (Story et al., 2003; Bandell et al., 2004; Bautista et al., 2005; Kobayashi et al., 2005). When overexpressed in cells, TRPA1 is activated by lower temperatures compared with TRPM8 (Story et al., 2003), but its role in cold transduction remains controversial (Bautista et al., 2006; Kwan et al., 2006). Previous studies suggested that TRPA1 is activated by elevation of [Ca2+]i during cooling rather than directly by cold (Zurborg et al., 2007). Indeed, Ca2+, possibly released from intracellular stores, can directly activate TRPA1 (Jordt et al., 2004; Doerner et al., 2007).
TRPA1 is activated by of mustard oil isothiocyanates (MO) and by other pungent natural products, including allicin, cinnamaldehyde, methylsalicylate, eugenol, and gingerol, as well as by environmental irritants, such as unsaturated aldehydes present in smoke or produced by drug metabolism, all of which elicit a painful burning or pricking sensation (Bandell et al., 2004; Jordt et al., 2004; Bautista et al., 2005; Xu et al., 2006). TRPV1 and TRPM8 instead do not respond to MO. Studies in mice lacking TRPA1 confirmed its role in nociception to irritants (Bautista et al., 2006; Kwan et al., 2006). Intrathecal injection of TRPA1-specific antisense oligonucleotides reverses the hyperalgesia induced by inflammation and nerve injury (Obata et al., 2005). Many TRPA1-activating compounds, like the alkylating agent iodoacetamide, form covalent adducts with thiols and primary amines (Hinman et al., 2006; Macpherson et al., 2007). Topical application of these agents excites sensory nerve fibers, producing acute pain and neurogenic inflammation through peripheral release of substance P, CGRP, purines, and other transmitters. It is noteworthy that MO, like capsaicin with TRPV1, causes functional homologous desensitization of TRPA1 (Ruparel et al., 2008).
MO also induces endothelium-independent and TRPA1-mediated vasorelaxation (Wilson et al., 2002). The plant-derived cannabinoid receptor agonists Δ9-tetrahydrocannabinol (THC) and cannabinol relax hepatic or mesenteric arteries in vitro by activating CGRP-containing sensory nerve endings that innervate the smooth muscle. This effect is not inhibited by antagonists of cannabinoid receptors, but it is blocked by ruthenium red, is extracellular Ca2+-dependent, and persists in TRPV1-deficient mice (Zygmunt et al., 2002). Experiments in TG neurons showed that a subpopulation of capsaicin-sensitive cells is responsive to both MO and THC, suggesting that nociceptors are excited through the same molecular mechanism, involving activation of a Ca2+-permeable channel on capsaicin-sensitive sensory neurons. In cells expressing TRPA1, THC or cannabinol activate this channel (Jordt et al., 2004), thus confirming that TRPA1 might contribute to inflammatory hypersensitivity and vasodilation.
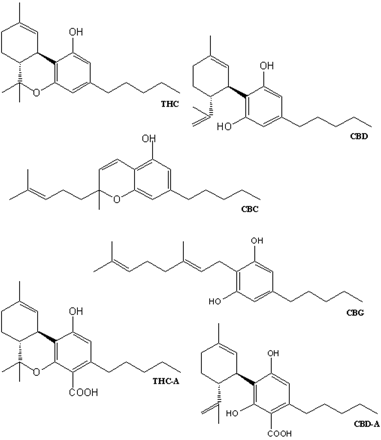
In view of the reported effects of phytocannabinoids on TRP channels (Bisogno et al., 2001; Jordt et al., 2004; Ligresti et al., 2006) and of the proposed therapeutic applications of cannabis extracts (Di Marzo and De Petrocellis, 2006), the aim of present work was to evaluate whether some of these natural products (Fig. 1) influence the activity of heterologously or constitutively expressed TRPA1 and TRPM8 channels on intracellular Ca2+ in HEK-293 cells and DRG neurons.
Materials and Methods
Drugs. All of the purified phytocannabinoids used were a kind gift from GW Pharmaceutical (Salisbury, Wilthshire, UK). Allyl isothiocyanate from mustard oil, trans-cinnamaldehyde, (-)menthol, and icilin were obtained from Sigma-Aldrich (St. Louis, MO). Ruthenium red was obtained from Alexis Biochemicals Axxora (San Diego, CA), and anandamide and Capsazepine were purchased from Tocris Cookson Ltd. (Bristol, UK). HU-210 was a gift from Prof. R. Mechoulam (Hebrew University of Jerusalem, Israel). All drugs were kept at -20°C, dry, and under an atmosphere of nitrogen until use. Under these conditions they are stable for at least 12 months. Before use, they were dissolved in dimethyl sulfoxide, whose final concentration in the assay buffer was never higher than 0.2%.
Stable Transfection of HEK-293 Cells with Rat TRPA1 cDNA and Rat TRPM8 cDNA Containing Plasmids. HEK-293 (human embryonic kidney) cells were grown as monolayers in minimum essential medium supplemented with nonessential amino acids, 10% fetal calf serum, and 2 mM glutamine, maintained under 5% CO2 at 37°C plated on 100-mm diameter Petri dishes, and transfected at approximately 80% confluence with Lipofectamine 2000 (Invitrogen, Carlsbad, CA) by using a plasmid pcDNA3 (Invitrogen) containing the rat TRPA1-cDNA or the rat TRPM8-cDNA (both are kind gift from Profs. Sven-Eric Jordt and David Julius, Department Cellular and Molecular Pharmacology, University of California, San Francisco, CA), according to the manufacturer’s protocol. Stably transfected clones were selected by G-418 (Geneticin; 600 μg/ml). Stable transfections were checked by quantitative real-time polymerase chain reaction (PCR).
Evaluation of TRPM8 and TRPA1 Receptor Transfection Levels by Quantitative Real-Time PCR. Total RNA was extracted by TRIzol (Invitrogen) according to the manufacturer’s recommendations, dissolved in RNA-storage solution (Ambion, Austin, TX), UV-quantified by a BioPhotometer (Eppendorf, New York, NY), and stored at -80°C. RNA aliquots (6 μg) were digested by RNase-free DNase I (Ambion; DNA-free kit) in a 20-μl final volume reaction mixture, to remove contaminating genomic DNA. After DNase digestion, concentration and purity of RNA samples were evaluated by the RNA-6000-Nano microchip assay, using a 2100 Bioanalyzer equipped with 2100 Expert Software (Agilent Technologies, Palo Alto, CA), following the manufacturer’s instructions. One microgram of total RNA, as evaluated by the 2100 Bioanalyzer, was reverse transcribed by MoMuLV Superscript III reverse transcriptase (Invitrogen), as described previously. Negative controls (-real-time) were produced by omitting the reverse transcriptase. Quantitative real-time PCR was performed by an iCycler-iQ (Bio-Rad, Hercules, CA) in a 25-μl reaction mixture containing the following: 1× iQ SYBR Green Supermix (Bio-Rad), 10 ng of cDNA (calculated on the basis of the retro-transcribed RNA), and 330 nM for each primer. The amplification profile consisted of an initial denaturation of 2 min at 94°C and 40 cycles of 30 s at 94°C, annealing for 30 s at TaOpt (optimum annealing temperature, see below), and elongation for 45 s at 68°C. Fluorescence data were collected during the elongation step. A final extension of 7 min was carried out at 72°C, followed by melt-curve data analysis. Optimized primers for SYBR Green analysis (and relative TaOpt) were designed by the Beacon-Designer software 6.0 version (Biosoft International, Palo Alto, CA) and were synthesized (HPLC-purification grade) by MWG-Biotech AG (Ebersberg, Germany). Assays were performed in quadruplicate (maximal ΔCt of replicate samples, <0.5), and a standard curve from consecutive 5-fold dilutions (100–0.16 ng) of the cDNAs was included, for PCR-efficiency determination. Relative expression analysis, correct for PCR efficiency and normalized with respect to reference genes β-actin and glyceraldehyde 3-phosphate dehydrogenase, was performed by GENEX software (Bio-Rad) for group-wise comparison and statistical analysis.
Experiments in HEK-293 Cells Overexpressing the Rat TRPA1 or TRPM8 Channels. Cells were plated on 100-mm diameter Petri dishes and after 3 days were loaded for 1 h at room temperature with the cytoplasmic calcium indicator Fluo4-AM (4 μM; Invitrogen) containing Pluronic (0.02%; Invitrogen). Cells were washed twice in Tyrode’s buffer (145 mM NaCl, 2.5 mM KCl, 1.5 mM CaCl2, 1.2 mM MgCl2, 10 mM d-glucose, and 10 mM HEPES, pH 7.4), resuspended in Tyrode’s buffer, and transferred to the quartz cuvette of the spectrofluorimeter (λex = 488 nm; λem = 516 nm) (Perkin-Elmer LS50B; PerkinElmer Life and Analytical Sciences, Waltham, MA) under continuous stirring. [Ca2+]i was determined before and after the addition of various concentrations of test compounds. EC50 values were determined as the concentration of test substances required to produce half-maximal increases in [Ca2+]i. Curve fitting and parameter estimation were performed with GraphPad Prism (GraphPad Software Inc., San Diego, CA). Phytocannabinoids were added 5 min before stimulation of cells with icilin or menthol. IC50 values were determined again using GraphPad Prism, because the concentration exerting a half-maximal inhibition of agonist induced increases of [Ca2+]i.
The efficacy of the agonists was first determined by normalizing their effect to the maximal effect on [Ca2+]i observed with 4 μM ionomycin. Subsequently, the effects of TRPA1 agonists are expressed as a percentage of the effect obtained with 100 μM MO. In experiments in TRPM8-HEK-293 cells, varying doses of phytocannabinoids were added 5 min (or in some experiments 15 min) before EC90 concentrations of menthol (50 μM) or icilin (0.25 μM). Data were expressed as the concentration exerting a half-maximal inhibition of agonist effect (IC50), which was calculated using GraphPad software. The effect on [Ca2+]i exerted by the agonist alone was taken as 100%.
Statistics. Statistical analysis of the data was performed by analysis of variance (ANOVA) at each point followed by the Bonferroni test. Differences were considered significant at P < 0.05.
Animals and Cells. DRG were prepared as described previously. DRG neurones from neonatal Sprague-Dawley rats (P7-P14) or adult mice were cultured as described previously (Vellani et al., 2001, 2004). In brief, isolated ganglia were digested in collagenase (type IV; Worthington Biochemical, Lakewood, NJ), mechanically dissociated, plated on glass coverslips coated with poly-l-lysine and laminin (Sigma-Aldrich), and maintained in Dulbecco’s modified Eagle’s medium (Invitrogen) containing 100 ng/ml nerve growth factor (100 ng/ml-1; Promega, Madison, WI), 10% fetal bovine serum, penicillin, and streptomycin. All animal procedures were carried out in accordance with Italian legislation. Neonatal rats (P7-P14) were killed by decapitation under halothane anesthesia, DRG were isolated, and neurons were dissociated and cultured. For dose-response experiments, approximately 10,000 to 30,000 neurons were used for each data point. Cells were exposed to capsaicin, mustard oil, and ionomycin; all of these treatments, which produce massive calcium influx from extracellular space, caused an increase in calcium in the cytoplasm.
Calcium Imaging in DRG Cultures. Calcium measurements were performed after 1 to 3 days in culture, DRG neurons were loaded in a serum-free medium for 10 min with the calcium-sensitive fluorescent indicator Fluo-4 AM (10 μM; Invitrogen). Experiments were carried out in the following extracellular solution: 145 mM NaCl, 4 mM KCl, 1.8 mM CaCl2, 1 mM MgCl2, 10 mM HEPES, and 4 mM glucose, neutralized to pH 7.4 with NaOH. Coverslips were mounted onto an upright E600 microscope (Nikon, Florence, IT) equipped with an Andor Technology imaging system (Belfast, Northern Ireland, UK). Fluo-4 AM was excited with 488-nm light, which was generated by a monochromator (Cairn Research, Faversham, UK), and images of the fluorescence (F) were captured every 2 s using a time course software package (Andor iQ; Andor Technology). The fluorescent images were acquired by an electron multiplying CCD camera (Andor Technology), and the fluorescence intensity in specific regions of interest was measured. Neurons were distinguished from non-neuronal cells at the start or at the end of the experiment by applying 25 mM KCl, which induce a rapid increase of [Ca2+]i in neurons. To identify neurons responding to both phytocannabinoids and allyl isothiocyanate or icilin, phytocannabinoids were added to DRG cultures at 100 μM concentration, 10 min before 100 μM MO or 1 μM icilin, and [Ca2+]i fluorescence was recorded for 180 s after the addition of each agonist. At the end of the experiment, the maximal fluorescence (Fmax) was obtained by application of the calcium ionophore ionomycin (10 μM; Calbiochem, La Jolla, CA) in the presence of high Ca2+ (30 mM) and K+ (125 mM). Calcium imaging data are expressed as F/Fmax. All experiments were performed at room temperature (23–25°C). In general, DRG neurons from neonatal rats were preferred for ease of supply and culture preparation. The main findings in DRG neurons isolated from neonatal rats were reproduced in cells from adult mice (data not shown), suggesting that the effects seen were neither species- nor age-dependent.
Morphology of DRG Neurons. DRG neuronal size was measured from calibrated images recorded in transmitted light before calcium imaging experiments. The area of individual neurons responding to specific agonists was measured by calculating the area of a corresponding region of interest drawn with Andor iQ software. Distribution histograms were generated using SigmaPlot 8.0 computer software (Systat Software Inc., San Jose, CA).
Statistics. The data are presented as mean ± S.E.M. values. Statistical analyses were performed using one-way ANOVA. Follow-up analysis was performed using the Bonferroni test. Total cell count and distribution of cell size were compared using the ANOVA comparison of log-transformed distribution data.
Results
HEK-293 Cell Transfection. We have stably transfected HEK-293 cells with the plasmid pcDNA3 containing the rat recombinant TRPA1 cDNA, thus generating cells (denoted with “TRPA1-HEK-293”) that, as assessed by real-time PCR, express high levels of TRPA1 transcript. This transcript was absent in HEK-293 cells stably transfected with cDNA encoding rat TRPM8 (denoted with “TRPM8-HEK-293”), which instead express high levels of TRPM8 transcript (De Petrocellis et al., 2007), and in HEK-293 cells that had not been transfected with the pcDNA3 plasmid and were treated with only Lipofectamine (data not shown).
TRPA1 Is Activated by Cinnamaldehyde and Other Sensory Compounds. Using a fluorometric test, we showed that rat TRPA1-HEK293 cells exhibit a sharp increase in intracellular [Ca2+]i upon application of MO or cinnamaldehyde, two compounds known to activate TRPA1. Using this test, we determined the concentration for half-maximal activation to be 13.0 ± 4.1 μM for cinnamaldehyde and 2.5 ± 0.7 μM for MO (Fig. 2A). Ruthenium red, a known blocker of TRPA1, inhibited the effect (data not shown). Cinnamaldehyde and MO did not activate TRPV1-HEK-293 or TRPM8-HEK-293 cells (data not shown). Analogous to what was reported by Jordt et al. (2004), THC also increased [Ca2+]i with an EC50 of 0.23 ± 0.03 μM (Fig. 2A).
TRPA1 Is Activated by Phytocannabinoids. All of the phytocannabinoids that were tested increased [Ca2+]i in TRPA1-HEK-293 cells. The EC50 values were (Hill slopes are between parentheses) as follows: cannabichromene (CBC), 0.06 ± 0.02 μM (1.1 ± 0.3); CBD, 0.096 ± 0.012 μM (0.8 ± 0.1); THC acid, 0.24 ± 0.03 μM (1.9 ± 0.5); CBD acid, 12.0 ± 8.8 μM (0.5 ± 0.1); cannabigerol (CBG), 3.4 ± 1.0 μM (0.6 ± 0.1) (means ± S.E., n = 4; Fig. 3B). In some cases, the response at 100 μM resulted higher than that of MO (THC = 117 ± 12%; CBC = 117 ± 10%; and CBG = 126 ± 11%), whereas the acids (CBD acid = 74 ± 6% and THC acid = 78 ± 8% of MO) showed a weaker effect. Therefore, CBD acid and THC acid behaved as partial agonists in this assay compared with MO, and this might be used for the future development of TRPA1 antagonists. Among the phytocannabinoids, only CBD increased [Ca2+]i also when tested in nontransfected HEK-293 cells, with a half-maximal activation value of 0.7 ± 0.2 μM but with a very low efficacy of 26 ± 3% at 50 μM. Therefore, the curves used to calculate its potency and efficacy at TRPA1 were subtracted from the corresponding values obtained in nontransfected cells. The synthetic cannabinoid receptor agonist, HU-210, did not exert any effect on TRPA1-HEK-293 cells up to a 10 μM concentration (data not shown). Note that 5-min preincubation of TRPA1-HEK-293 cells with CBC or CBG prevented the elevation of [Ca2+]i induced by MO in TRPA1-HEK-293 cells, with IC50 values of 0.09 ± 0.02 and 1.0 ± 0.01 μM, respectively (Fig. 2C).
 Fig. 4.
Fig. 4.
Dose-related inhibition by various phytocannabinoids (THC, CBD, CBG, THC acid, and CBD acid) on icilin (A) and menthol (B)-induced elevation of [Ca2+]i in HEK-293 cells stably transfected with the rat recombinant TRPM8 channel. Icilin and menthol elicited no response in nontransfected cells (see Results). Data are expressed as percentage of the maximal effect observed with either icilin (0.25 μM) (A) or menthol (50 μM) (B). The effect observed for each dose of CBD was corrected for its stimulatory effect on [Ca2+]i observed in nontransfected cells. Data are the means ± S.E. of n = 4 separate determinations. S.E. bars are not shown for the sake of clarity and were never higher than 5% of the means. In both A and B, the effect of concentrations ≥10-7 M of all compounds, with the exception of CBC, and of concentrations ≥10-6 M for CBD acid was statistically significant (P < 0.01, as assessed by ANOVA followed by the Bonferroni test).
Effect of Phytocannabinoids on Icilin-Responding DRG Neurons. Because THC acid and THC are quite efficacious at TRPA1 (present data) and CBD also activates TRPV1, we used CBG, which was the least potent compound on TRPA1 and one of the most potent ones at antagonizing TRPM8-induced elevation of [Ca2+]i in TRPM8-HEK-293 cells (present data), to study the effect on DRG neurons. Although CBG (100 μM) also elevated [Ca2+]i in neurons that did not respond to icilin (Fig. 5A), possibly due to interaction with other TRP channels (notably CBG, apart from TRPA1, also activates TRPV1 at high concentrations), it did inhibit the response to icilin in icilin-responding DRG neurons in a dose-dependent way (IC50 = 4.5 μM, 5-min preincubation; Fig. 5B). Note that, anandamide, which was previously found to inhibit menthol- and icilin-induced response in TRPM8-HEK-293 cells (De Petrocellis et al., 2007), also inhibited the icilin response in DRG neurons with a potency similar to that of CBG (Fig. 5C). The inhibitory effect of CBG was unlikely due to its elevation of [Ca2+]i, and subsequent TRPM8 desensitization, for three reasons: 1) the size distribution of icilin-responding neurons was different from the size distribution of CBG-responding cells, the former being composed exclusively of small, and the latter of medium to large, neurons, with very little overlap (Fig. 5A); 2) almost no individual neuron responding to icilin responded to CBG (100 μM) with further elevation of [Ca2+]i, again suggesting that CBG-induced calcium response occurred only in non-TRMP8-expressing neurons; and 3) when it inhibited the icilin response, CBG was inactive at elevating [Ca2+]i.

Effect of CBG on basal and icilin (1 μM)-induced elevation of [Ca2+]i in neonatal DRG neurons as assessed by calcium imaging. A, size distribution of the entire DRG neuron population of neurons responding to either icilin (1 μM) (black bars) or CBG (100 μM) (white bars) with increased [Ca2+]i. B, dose-related effect of CBG on icilin-induced elevation of [Ca2+]i. C, dose-related effect of anandamide on icilin-induced elevation of [Ca2+]i. Data are the means ± S.E. of n = 10,000 to 30,000 neurons.
Effects of CB1 Receptor Antagonists on Phytocannabinoid-Induced Responses. Because (especially in DRG neurons) some of the effects of the phytocannabinoids were observed at concentrations higher than 10 μM, the possibility exists that these effects are mediated by interaction with CB1 receptors, which are expressed in these sensory neurons as well as, to some extent, in HEK-293 cells (De Petrocellis et al., 2007). Instead, CB2 receptors are not present in HEK-293 cells and DRG neurons. For this reason, we tested the effect of CB1 receptor antagonists on the effects of CBC on TRPA1 and CBG on TRPM8. In TRPA1-HEK-293 cells, AM251 and LY320135 reduced the MO (100 μM)-induced effect on [Ca2+]i at concentrations ≥1 μM and, therefore, were tested on CBC at a concentration of 0.5 μM, which are well above their Ki for CB1 receptors (Gatley et al., 1997; Felder et al., 1998). AM251 nonsignificantly reduced (P > 0.05) the effect of CBC (10 μM) by 24 ± 9% (mean ± S.D., n = 3), whereas LY320135 exerted no effect (4 ± 1% inhibition, mean ± S.D., n = 3). In TRPM8-HEK-293 cells, AM251, as previously shown for SR141716A (De Petrocellis et al., 2007), potently inhibited icilin (0.25 μM)-induced elevation of [Ca2+]i at concentrations (0.1–0.2 μM and above) similar to those required to antagonize CB1, whereas LY320135 was inactive at concentrations <0.5 μM. Therefore, we only tested this compound on CBG (10 μM) inhibition of icilin-induced elevation of [Ca2+]i. LY320135 (0.1 and 0.25 μM) exerted no statistically significant inhibition of CBG effect (+6 ± 2 and -15 ± 8%, P > 0.05, means ± S.D., n = 3).
In cultured DRG cells, LY320135 at concentrations up to 10 μM per se did not produce any effect on intracellular calcium concentration. A total of 100 μM CBC produced a Ca2+ rise in 29.6 ± 3.5% of cultured DRG neurons and in 25.6 ± 2.6% when 1 μM LY320135 was added 2 to 4 min before CBC addition; this difference was not statistically significant (data were from three separate cultures). Likewise, when 10 μM LY320135 was present, the effect of 100 μM CBC was seen in 22.5 ± 1.5% of cultured DRG neurons, and, again, the difference was not statistically significant. We then applied solutions containing a mixture of CBG, LY320135, and icilin to assess whether the inhibitory effect of CBG on icilin-induced Ca2+ elevation was due to CB1 receptors. However, the combination of such compounds, even if icilin was applied after preapplication of a mixture of CBG (100 μM) and LY320135 (1 or 10 μM), produced large calcium responses in a significant number of DRG non-neuronal cells present in the culture and in many large-sized DRG neurones, which normally do not respond to icilin (see Fig. 5). Due to this aberrant effect of LY320135/CBG/icilin, the role of CB1 receptors in CBG modulation of icilin response could not be fully investigated in DRG neurons.
Discussion
In this study, we have described for the first time the capability of several phytocannabinoids to influence the TRPA1- and TRPM8-mediated elevation of [Ca2+]i in HEK-293 cells overexpressing these channels and in DRG sensory neurons. Because elevation of [Ca2+]i plays a fundamental role in the function of these channels, which is not limited to induce excitation and depolarization of sensory neurons and subsequent thermoreception, but might also play a role in the survival of some cancer cells (Zhang and Barritt 2004), as well as in the mobilization of intracellular calcium and subsequent activation of Ca2+-operated channels (Thebault et al., 2005; Mahieu et al., 2007) and the sensitization/desensitization of other channels including many TRPs (Levine and Alessandri-Haber, 2007), these data raise the possibility that the studied phytocannabinoids might exert some of their several pharmacological activities by also regulating TRPA1 and TRPM8 activity. Phytocannabinoids, and CBC in particular, potently and efficaciously elevated [Ca2+]i in TRPA1-expressing, but not wild-type, HEK-293 cells and in MO-responding DRG neurons. The potency of CBC in DRG neurons (∼34 μM) was, as expected when using cells that constitutively express receptors, considerably lower than that in HEK-293 cells (61 nM), although the efficacy was comparable with that of MO in both cell types. It is interesting to note that the rank of potency in TRPA1-HEK-293 cells (CBC>CBD>THC>THC-acid>CBG >CBD-acid) reflects the expected electrophilic nature of the hydroxy group in position 1. This is highest in CBC, in which a partly unsaturated, electron-attracting pyrane group is conjugated with the aromatic cycle, and lowest in THC acid, CBD acid, and CBG, in which the pyrane moiety is absent and the strong electron donor groups are present. However, the only exception to this rule is CBD, which might exert a more potent effect due to its ability to elevate [Ca2+]i also in a TRPA1-independent manner and to the well known property of Ca2+ to directly activate TRPA1. Therefore, given the well established fact that TRPA1 are activated by compounds containing electrophilic groups, which seem to form covalent bonds with cysteine groups of the protein, thereby gating the passage of calcium (Hinman et al., 2006; Macpherson et al., 2007), these observations suggest that phytocannabinoids might also activate the channel in this way. However, specific studies, including a direct demonstration of the interaction between, e.g., CBC and TRPA1 obtained by means of electrophysiological (i.e., patch-clamp) measurements and site-directed mutagenesis of the cDNA encoding this protein, need to be carried out to demonstrate that this hypothesis is correct.
Although in DRG neurons a large superimposition between the size of cells responding to MO and that of the cells responding to CBC was observed, the kinetics of [Ca2+]i elevation induced by the phytocannabinoid was different from that observed in TRPA1-HEK-293 cells. This phenomenon, like the lower potency observed in these neurons, might be due to the lower degree of expression of TRPA1 in sensory neurons compared with HEK-293 cells stably transfected with TRPA1 cDNA, as well as to the presence of other channels that might respond to CBC. These channels are unlikely to be TRPV1 channels (which are highly coexpressed in TRPA1-expressing neurons), because we have previously found that both human and rat recombinant TRPV1 is insensitive to CBC (Ligresti et al., 2006). However, other channels, including but not limited to TRPV channels, might also be activated by this compound and influence the kinetics of the calcium response in sensory neurons. It is unfortunate that, to date, the lack of specific TRPA1 antagonists did not allow us to rule out the possibility that other molecular targets might contribute to the CBC effect on [Ca2+]i in these neurons. At any rate, in view of the vasodilatory and nociceptive responses caused by TRPA1 activation (Bautista et al., 2005), and in view of their easy desensitization by TRPA1 agonists (Ruparel et al., 2008), our findings suggest that the anti-inflammatory and antinociceptive effects of cannabis extracts and CBD might be due, in part, to the capability of phytocannabinoids to activate/desensitize TRPA1. In this study, we have found that preincubation with CBC and CBG prevents the elevation of [Ca2+]i in TRPA1-HEK-293 cells and DRG neurons. Accordingly, during the revision process of the present article, a study reported that, in trigeminal sensory neurons, some synthetic cannabinoids, by activating TRPA1 at concentrations similar to those reported here for phytocannabinoids in DRG neurons, desensitize MO-induced and TRPA1-mediated currents (Akopian et al., 2008).
We have also shown here that although TRPM8 is not activated by phytocannabinoids, its effect on [Ca2+]i elevation, induced by two different agents (menthol and icilin), is inhibited by some of these compounds. This finding was not particularly unexpected because we have previously shown that the endocannabinoids, anandamide and N-arachidonoyldopamine, antagonize the TRPM8-mediated response to menthol and icilin (De Petrocellis et al., 2007), and several authors have reported that agents that activate TRPV1 (and CBD and CBG are listed among these) usually produce the opposite effect on TRPM8 channels (see Introduction). Here, we found that all of the phytocannabinoids tested, including THC, and with the exception of CBD acid (which was a relatively weak antagonist) and CBC (which was completely inactive), were nearly equipotent at antagonizing the TRPM8-mediated response to icilin and menthol. TRPM8 antagonism by CBG was also found to occur in DRG neurons, and, in this case, although this compound exerted a stimulatory effect on some non-icilin-responding neurons of medium-to-large diameter per se, in those small neurons that did respond to icilin, the potency of the inhibitory effect on [Ca2+]i elevation (IC50 ∼4.5 μM) was not exceedingly lower than that observed in TRPM8-HEK-293 cells (IC50 ∼0.14 μM). Although the role of TRPM8 in nociception (other than that induced by cooling) is still controversial, this channel has been strongly associated with cancer cell proliferation, particularly in prostate carcinoma (Tsavaler et al., 2001; Thebault et al., 2005). In fact, it has been shown that the survival of prostate cancer cells depends on the presence of TRPM8 channels, and hence TRPM8 antagonists have been suggested as a potential therapy for the treatment of this type of cancer (Zhang and Barritt 2004; Bidaux et al., 2007). Specific studies in this direction need to be performed before it can be suggested that phytocannabinoids with antagonist activity at TRPM8 can be used as anticancer agents. Furthermore, the nature of the interaction between phytocannabinoids and TRPM8 needs to be investigated in detail, for example, through the use of electrophysiological (i.e., patch-clamp) measurements. In fact, it is possible that these compounds influence the capability of icilin and menthol to gate TRPM8 channels not just because they are interacting with their binding sites on the protein, but also because they cause effects downstream of TRPM8 gating.
In DRG neurons, where, as stated above, the effects described here were observed at concentrations of phytocannabinoids generally higher than 1 μM, it is possible that these lipophilic compounds also act either by membrane bilayer perturbation or by activating cannabinoid CB1 receptors, which are known to be expressed in sensory neurons and, to some extent, also in HEK-293 cells (De Petrocellis et al., 2007). However, although it has been suggested that lipophilicity is a “component, but not a primary determinant of pharmacological activity in the cannabinoids” (Thomas et al., 1990), we found that in both DRG neurons and HEK-293 cells, CB1 receptor antagonists do not significantly affect the ability of CBC to activate TRPA1-mediated elevation of intracellular Ca2+, and that at least in HEK-293 cells also, the inhibitory effect of CBG on icilin-induced responses was not influenced by one of these antagonists. Accordingly, Akopian et al. (2008) showed that two very potent CB1 and/or CB2 agonists activate TRPA1 in trigeminal neurons in a way completely independent from cannabinoid receptors.
In conclusion, we have reported here for the first time the potent and efficacious modulatory effects by some phytocannabinoids on TRPA1- and TRPM8-mediated [Ca2+]i elevation, and in particular the stimulation of CBC on TRPA1 activity, and the inhibition by THC, THC acid, CBD, and CBG of TRPM8 activity in HEK-293 cells stably transfected with the cDNAs encoding for the two proteins. Similar effects were also found to occur in DRG sensory neurons. The exact molecular nature of these interactions, as well as their potential therapeutic relevance, will have to be addressed in future studies. Nevertheless, in view of the following observations: 1) TRPA1 and TRPM8 participate in pain transduction (Obata et al., 2005); 2) TRPA1 stimulation by synthetic cannabinoids causes its desensitization and is responsible for some of the analgesic effects of these compounds (Akopian et al., 2008; Ruparel et al., 2008); 3) the phytocannabinoids tested here can represent up to 0.06 (CBC), 0.62 (CBG), 0.86 (CBD), 9.7 (CBD acid), 13.0 (THC acid), and 1.34 (THC) percent in weight of dried cannabis flowers (personal communication from Dr. P. Gibson and A. Sutton, GW Pharmaceuticals); and 4) the rat TRPA1 and TRPM8 (used in the present study) compared to their human orthologs (GenBank protein sequences: TRPA1 Rattus norvegicus NP_997491; TRPA1 Homo sapiens NP_015628; TRPM8 R. norvegicus NP_599198; and TRPM8 H. sapiens NP_076985) exhibit extremely similar amino acid sequences (89% for TRPA1 and 97% for TRPM8) and analogous predicted three-dimensional structure; our findings suggest that some of the analgesic effects of cannabis extracts reported in clinical trials (for review, see Di Marzo and De Petrocellis, 2006) might be due to interaction with TRPA1 and TRPM8 channels.
Footnotes
-
This work was supported by grants from GW Pharmaceuticals, UK (to V.D.M.), the Italian Ministry of Education, University and Scientific Research (PRIN 2004057339_002) (to V.V.), and Fondazione Cassa di Risparmio di Modena and Fondazione Cassa di Risparmio di Carpi (to V.V.).
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.134809.
-
ABBREVIATIONS: TRP, transient receptor potential; TRPV, TRP channels of vanilloid; TRPA, transient receptor potential channel of ankyrin; DRG, dorsal root ganglia; TG, trigeminal; TRPV1, TRP type 1; CBD, cannabidiol; TRPA1, TRP channel of ankyrin type; TRPM8, TRP channel of melastatin type 8; CGRP, calcitonin gene-related peptide; MO, mustard oil isothiocyanates; THC, Δ9-tetrahydrocannabinol; anandamide, N-arachidonoylethanolamine; Capsazepine, N-[2-(4-chlorophenyl)ethyl]-1,3,4,5-tetrahydro-7,8-dihydroxy-2H-2-benzazepine-2-carbothioamide; PCR, polymerase chain reaction; ANOVA, analysis of variance; F, fluorescence; Fmax, maximal fluorescence; CBC, cannabichromene; CBG, cannabigerol; AM251, N-(piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide; LY320135, 6-methoxy-2-(4-methoxyphenyl)benzo[b]-thien-3-yl][4-cyanophenyl]methanone; SR141716A, N-(piperidin-1-yl)-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide; HU-210, (6a,R)-trans-3-(1,1-dimethylheptyl)-6a,7,10,10a-tetrahydro-1-hydroxy-6,6-dimethyl-6H-dibenzo[b,d]pyran-9-methanol.
- Received November 29, 2007.
- Accepted March 18, 2008.
- The American Society for Pharmacology and Experimental Therapeutics