Epipelagic (talk | contribs) m →Deep ocean webs: ~ |
add attribution for licensed material |
||
| Line 378: | Line 378: | ||
| image1 = Importance of Antarctic krill in biogeochemical cycles.png |
| image1 = Importance of Antarctic krill in biogeochemical cycles.png |
||
| caption1 = {{center|'''Processes in the biological pump'''}}{{center|<small> Numbers given are carbon fluxes (Gt C yr−1) in white boxes<br />and carbon masses (Gt C) in dark boxes</small>}} |
| caption1 = {{center|'''Processes in the biological pump'''}}{{center|<small> Numbers given are carbon fluxes (Gt C yr−1) in white boxes<br />and carbon masses (Gt C) in dark boxes</small>}} |
||
Phytoplankton convert CO2, which has dissolved from the atmosphere into the surface oceans into particulate organic carbon (POC) during primary production. Phytoplankton are then consumed by krill and small zooplankton grazers, which in turn are preyed upon by higher trophic levels. Any unconsumed phytoplankton form aggregates, and along with zooplankton faecal pellets, sink rapidly and are exported out of the mixed layer. Krill, zooplankton and microbes intercept phytoplankton in the surface ocean and sinking detrital particles at depth, consuming and respiring this POC to CO2 (dissolved inorganic carbon, DIC), such that only a small proportion of surface-produced carbon sinks to the deep ocean (i.e., depths > 1000 m). As krill and smaller zooplankton feed, they also physically fragment particles into small, slower- or non-sinking pieces (via sloppy feeding, coprorhexy if fragmenting faeces), retarding POC export. This releases dissolved organic carbon (DOC) either directly from cells or indirectly via bacterial solubilisation (yellow circle around DOC). Bacteria can then remineralise the DOC to DIC (CO2, microbial gardening). Diel vertically migrating krill, smaller zooplankton and fish can actively transport carbon to depth by consuming POC in the surface layer at night, and metabolising it at their daytime, mesopelagic residence depths. Depending on species life history, active transport may occur on a seasonal basis as well.<ref>Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". ''Nature communications'', '''10'''(1): 1–13. {{doi|10.1038/s41467-019-12668-7}}.</ref> |
Phytoplankton convert CO2, which has dissolved from the atmosphere into the surface oceans into particulate organic carbon (POC) during primary production. Phytoplankton are then consumed by krill and small zooplankton grazers, which in turn are preyed upon by higher trophic levels. Any unconsumed phytoplankton form aggregates, and along with zooplankton faecal pellets, sink rapidly and are exported out of the mixed layer. Krill, zooplankton and microbes intercept phytoplankton in the surface ocean and sinking detrital particles at depth, consuming and respiring this POC to CO2 (dissolved inorganic carbon, DIC), such that only a small proportion of surface-produced carbon sinks to the deep ocean (i.e., depths > 1000 m). As krill and smaller zooplankton feed, they also physically fragment particles into small, slower- or non-sinking pieces (via sloppy feeding, coprorhexy if fragmenting faeces), retarding POC export. This releases dissolved organic carbon (DOC) either directly from cells or indirectly via bacterial solubilisation (yellow circle around DOC). Bacteria can then remineralise the DOC to DIC (CO2, microbial gardening). Diel vertically migrating krill, smaller zooplankton and fish can actively transport carbon to depth by consuming POC in the surface layer at night, and metabolising it at their daytime, mesopelagic residence depths. Depending on species life history, active transport may occur on a seasonal basis as well.<ref>Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". ''Nature communications'', '''10'''(1): 1–13. {{doi|10.1038/s41467-019-12668-7}}. [[File:CC-BY icon.svg|50px]] Material was copied from this source, which is available under a [https://creativecommons.org/licenses/by/4.0/ Creative Commons Attribution 4.0 International License].</ref> |
||
}} |
}} |
||
Revision as of 11:20, 28 March 2020

| Part of a series of overviews on |
| Marine life |
|---|
 |
Compared to terrestrial environments, marine environments have biomass pyramids which are inverted at the base. In particular, the biomass of consumers (copepods, krill, shrimp, forage fish) is larger than the biomass of primary producers. This happens because the ocean's primary producers are tiny phytoplankton which grow and reproduce rapidly, so a small mass can have a fast rate of primary production. In contrast, terrestrial primary producers, such as mature forests, grow and reproduce slowly, so a much larger mass is needed to achieve the same rate of primary production.
Because of this inversion, it is the zooplankton that make up most of the marine animal biomass. As primary consumers, zooplankton are the crucial link between the primary producers (mainly phytoplankton) and the rest of the marine food web (secondary consumers).[1]
If phytoplankton dies before it is eaten, it descends through the euphotic zone as part of the marine snow and settles into the depths of sea. In this way, phytoplankton sequester about 2 billion tons of carbon dioxide into the ocean each year, causing the ocean to become a sink of carbon dioxide holding about 90% of all sequestered carbon.[2] The ocean produces about half of the world's oxygen and stores 50 times more carbon dioxide than the atmosphere.[3]
An ecosystem cannot be understood without knowledge of how its food web determines the flow of materials and energy. Phytoplankton autotrophically produced biomass by converting inorganic compounds into organic ones. In this way, phytoplankton function as the foundation of the marine food web by supporting all other life in the ocean. The second central process in the marine food web is the microbial loop. This loop degrades marine bacteria and archaea, remineralises organic and inorganic matter, and then recycles the product either within the pelagic food web or by depositing it as sediment on the seafloor.[4]
Trophic levels
| marine food chain (typical) |
|---|
|
|
| ↓ |
| phytoplankton |
| ↓ |
| herbivorous zooplankton |
| ↓ |
| carnivorous zooplankton |
| ↓ |
|
|
| ↓ |
|
|
All life forms in the sea have the potential and perhaps the destiny to become food for something else. In the ocean, the food chain typically starts with phytoplankton and follows a course such as:
Phytoplankton → herbivorous zooplankton → carnivorous zooplankton → filter feeder → predatory vertebrate

Phytoplankton are the main primary producers at the bottom of the marine food chain. Since they are at the first level in the food chain they are said to have a trophic level of 1 (from the Greek trophē meaning food). Phytoplankton use photosynthesis to convert inorganic carbon into protoplasm. They are then consumed by microscopic animals called zooplankton.
Zooplankton comprise the second trophic level in the food chain, and include microscopic one-celled organisms called protozoa as well as small crustaceans, such as copepods and krill, and the larva of fish, squid, lobsters and crabs.
In turn, small zooplankton are consumed by both larger predatory zooplankters, such as krill, and by forage fish, which are small, schooling, filter-feeding fish. This makes up the third trophic level in the food chain.
The fourth trophic level consists of predatory fish, marine mammals and seabirds that consume forage fish. Examples are swordfish, seals and gannets.
Apex predators, such as orcas, which can consume seals, and shortfin mako sharks, which can consume swordfish, make up a fifth trophic level. Baleen whales can consume zooplankton and krill directly, leading to a food chain with only three or four trophic levels.
In practice, trophic levels are not always simple integers because the same consumer species often feeds across more than one trophic level.[6][7] For example a large marine vertebrate may eat smaller predatory fish but may also eat filter feeders; the stingray eats crustaceans, but the hammerhead eats both crustaceans and stingrays. Animals can also eat each other; the cod eats smaller cod as well as crayfish, and crayfish eat cod larvae. The feeding habits of a juvenile animal, and, as a consequence, its trophic level, can change as it grows up.
The fisheries scientist Daniel Pauly sets the values of trophic levels to one in plants and detritus, two in herbivores and detritivores (primary consumers), three in secondary consumers, and so on. The definition of the trophic level, TL, for any consumer species is:[8]

where is the fractional trophic level of the prey j, and represents the fraction of j in the diet of i. In the case of marine ecosystems, the trophic level of most fish and other marine consumers takes value between 2.0 and 5.0. The upper value, 5.0, is unusual, even for large fish,[9] though it occurs in apex predators of marine mammals, such as polar bears and killer whales.[10]


Primary producers
At the base of the ocean food web are single-celled algae and other plant-like organisms known as phytoplankton. Like plants on land, phytoplankton use chlorophyll and other light-harvesting pigments to carry out photosynthesis, absorbing atmospheric carbon dioxide to produce sugars for fuel. Chlorophyll in the water changes the way it reflects and absorbs sunlight, allowing scientists to map the amount and location of phytoplankton. These measurements give scientists valuable insights into the health of the ocean environment, and help scientists study the ocean carbon cycle.[11]


Among the phytoplankton are members from a phylum of bacteria called cyanobacteria. Marine cyanobacteria include the smallest known photosynthetic organisms. The smallest of all, Prochlorococcus, is just 0.5 to 0.8 micrometres across.[12] In terms of individual numbers, Prochlorococcus is possibly the most plentiful species on Earth: a single millilitre of surface seawater can contain 100,000 cells or more. Worldwide there are estimated to be several octillion (~1027) individuals.[13] Prochlorococcus is ubiquitous between 40°N and 40°S and dominates in the oligotrophic (nutrient poor) regions of the oceans.[14] The bacterium accounts for about 20% of the oxygen in the Earth's atmosphere.[15]
In oceans, most primary production is performed by algae. This is a contrast to on land, where most primary production is performed by vascular plants. Algae ranges from single floating cells to attached seaweeds, while vascular plants are represented in the ocean by groups such as the seagrasses. Larger producers, such as seagrasses and seaweeds, are mostly confined to the littoral zone and shallow waters, where they attach to the underlying substrate and are still within the photic zone. But most of the primary production by algae is performed by the phytoplankton.
- Phytoplankton form the base of the ocean foodchain
-
 Phytoplankton
Phytoplankton -
 Dinoflagellate
Dinoflagellate -
 Diatoms
Diatoms



Thus, in ocean environments, the first bottom trophic level is occupied principally by phytoplankton, microscopic drifting organisms, mostly one-celled algae, that float in the sea. Most phytoplankton are too small to be seen individually with the unaided eye. They can appear as a green discoloration of the water when they are present in high enough numbers. Since they increase their biomass mostly through photosynthesis they live in the sun-lit surface layer (euphotic zone) of the sea.
The most important groups of phytoplankton include the diatoms and dinoflagellates. Diatoms are especially important in oceans, where according to some estimates they contribute up to 45% of the total ocean's primary production.[16] Diatoms are usually microscopic, although some species can reach up to 2 millimetres in length.
Primary consumers
The second trophic level (primary consumers) is occupied by zooplankton which feed off the phytoplankton. Together with the phytoplankton, they form the base of the food pyramid that supports most of the world's great fishing grounds. Zooplankton are tiny animals found with the phytoplankton in oceanic surface waters, and include tiny crustaceans, and fish larvae and fry (recently hatched fish). Most zooplankton are filter feeders, and they use appendages to strain the phytoplankton in the water. Some larger zooplankton also feed on smaller zooplankton. Some zooplankton can jump about a bit to avoid predators, but they can't really swim. Like phytoplankton, they float with the currents, tides and winds instead. Zooplanktons can reproduce rapidly, their populations can increase up to thirty percent a day under favourable conditions. Many live short and productive lives and reach maturity quickly.
- Zooplankton form a second level in the ocean food chain
-
 Segmented worm
Segmented worm -
 Tiny shrimp-like crustaceans
Tiny shrimp-like crustaceans -
 Juvenile planktonic squid
Juvenile planktonic squid



Particularly important groups of zooplankton are the copepods and krill. These are not shown in the images above, but are discussed in more detail later. Copepods are a group of small crustaceans found in ocean and freshwater habitats. They are the biggest source of protein in the sea,[17] and are important prey for forage fish. Krill constitute the next biggest source of protein. Krill are particularly large predator zooplankton which feed on smaller zooplankton. This means they really belong to the third trophic level, secondary consumers, along with the forage fish.
Together, phytoplankton and zooplankton make up most of the plankton in the sea. Plankton is the term applied to any small drifting organisms that float in the sea (Greek planktos = wanderer or drifter). By definition, organisms classified as plankton are unable to swim against ocean currents; they cannot resist the ambient current and control their position. In ocean environments, the first two trophic levels are occupied mainly by plankton. Plankton are divided into producers and consumers. The producers are the phytoplankton (Greek phyton = plant) and the consumers, who eat the phytoplankton, are the zooplankton (Greek zoon = animal).

Traditionally jellyfish have been viewed as trophic dead ends, minor players in the marine food web, gelatinous organisms with a body plan largely based on water that offers little nutritional value for other organisms apart from a few specialised predators such as the ocean sunfish and the leatherback sea turtle.[19][18] That view has recently been challenged. Jellyfish, and more generally "gelatinous zooplankton" which include salps and ctenophores, are very diverse, fragile with no hard parts, difficult to see and monitor, subject to rapid population swings and often live inconveniently far from shore or deep in the ocean. It is difficult for scientists to detect and analyse jellyfish in the guts of predators, since they turn to mush when eaten and are rapidly digested.[19] But jellyfish bloom in vast numbers, and it has been shown they form major components in the diets of tuna, spearfish and swordfish as well as various birds and invertebrates such as octopus, sea cucumbers, crabs and amphipods.[20][18] "Despite their low energy density, the contribution of jellyfish to the energy budgets of predators may be much greater than assumed because of rapid digestion, low capture costs, availability, and selective feeding on the more energy-rich components. Feeding on jellyfish may make marine predators susceptible to ingestion of plastics."[18]
Higher order consumers
- Marine invertebrates
- Fish

Forage fish occupy central positions in the ocean food webs. The organisms it eats are at a lower trophic level, and the organisms that eat it are at a higher trophic level. Forage fish occupy middle levels in the food web, serving as a dominant prey to higher level fish, seabirds and mammals.
- Other marine vertebrates
In 2010 researchers found whales carry nutrients from the depths of the ocean back to the surface using a process they called the whale pump.[22] Whales feed at deeper levels in the ocean where krill is found, but return regularly to the surface to breathe. There whales defecate a liquid rich in nitrogen and iron. Instead of sinking, the liquid stays at the surface where phytoplankton consume it. In the Gulf of Maine the whale pump provides more nitrogen than the rivers.[23]
-
 Humpback whales lunge feeding on forage fish
Humpback whales lunge feeding on forage fish -



-
 Whale pump nutrient cycle
Whale pump nutrient cycle -
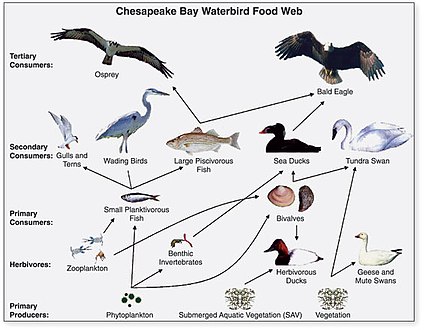 Waterbird food web in Chesapeake Bay
Waterbird food web in Chesapeake Bay


Microorganisms

There has been increasing recognition in recent years that marine microorganisms play much bigger roles in marine ecosystems than was previously thought. Developments in metagenomics gives researchers an ability to reveal previously hidden diversities of microscopic life, offering a powerful lens for viewing the microbial world and the potential to revolutionise understanding of the living world.[25] Metabarcoding dietary analysis techniques are being used to reconstruct food webs at higher levels of taxonomic resolution and are revealing deeper complexities in the web of interactions.[26]
Microorganisms play key roles in marine food webs. The viral shunt pathway is a mechanism that prevents marine microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms.[27] Viral shunting helps maintain diversity within the microbial ecosystem by preventing a single species of marine microbe from dominating the micro-environment.[28] The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.[29]

as imaged by a satellite in 2011



Fungi

- Amend, A., Burgaud, G., Cunliffe, M., Edgcomb, V.P., Ettinger, C.L., Gutiérrez, M.H., Heitman, J., Hom, E.F., Ianiri, G., Jones, A.C. and Kagami, M. (2019) "Fungi in the marine environment: Open questions and unsolved problems". MBio, 10(2): e01189-18. doi:10.1128/mBio.01189-18.
Coastal webs

- Byrnes, J.E., Reynolds, P.L. and Stachowicz, J.J. (2007) "Invasions and extinctions reshape coastal marine food webs". PloS one, 2(3): e295. doi:10.1371/journal.pone.0000295



DNA barcoding can be used to construct food web structures with better taxonomic resolution at the web nodes. This provides more specific species identification and greater clarity about exactly who eats whom. "DNA barcodes and DNA information may allow new approaches to the construction of larger interaction webs, and overcome some hurdles to achieving adequate sample size".[26]
Deep ocean webs
- Mesopelagic fishes
- Mesopelagic food web
-
![Impact of mesopelagic species on the global carbon budget [37] DVM = diel vertical migration NM = non-migration](https://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Mesopelagic_species_impact_on_global_carbon_budget.png/602px-Mesopelagic_species_impact_on_global_carbon_budget.png) Impact of mesopelagic species on the global carbon budget [37]
Impact of mesopelagic species on the global carbon budget [37]
DVM = diel vertical migration NM = non-migration
![Impact of mesopelagic species on the global carbon budget [37] DVM = diel vertical migration NM = non-migration](https://thcscience.wiki/term/cannabaceae/cannabis/cannabis-indica/?rdp_we_resource=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FFile%3AMesopelagic_species_impact_on_global_carbon_budget.png)
Scientists are starting to explore in more detail the largely unknown twilight zone of the mesopelagic, 200 to 1,000 metres deep. This layer is responsible for removing about 4 billion tonnes of carbon dioxide from the atmosphere each year. The mesopelagic layer is inhabited by most of the marine fish biomass.[38]

- Fish in the twilight cast new light on ocean ecosystem The Conversation, 10 February 2014.
- An Ocean Mystery in the Trillions The New York Times, 29 June 2015.
- Mesopelagic fishes - Malaspina circumnavigation expedition of 2010.[39][40]

According to a 2017 study, narcomedusae consume the greatest diversity of mesopelagic prey, followed by physonect siphonophores, ctenophores and cephalopods. The importance of the so called "jelly web" is only beginning to be understood, but it seems medusae, ctenophores and siphonophores can be key predators in deep pelagic food webs with ecological impacts similar to predator fish and squid. Traditionally gelatinous predators were thought ineffectual providers of marine trophic pathways, but they appear to have substantial and integral roles in deep pelagic food webs.[42] Diel vertical migration, an important active transport mechanism, allows mesozooplankton to sequester carbon dioxide from the atmosphere as well as supply carbon needs for other mesopelagic organisms.[43]
- Seeps and vents


Polar webs

surrounded by oceans

surrounded by landmasses
Arctic and Antarctic marine systems have very different topographical structures and as a consequence have very different food web structures.[48] Both Arctic and Antarctic pelagic food webs have characteristic energy flows controlled largely by a few key species. But there is no single generic web for either. Alternative pathways are important for resilience and maintaining energy flows. However, these more complicated alternatives provide less energy flow to upper trophic-level species. "Food-web structure may be similar in different regions, but the individual species that dominate mid-trophic levels vary across polar regions".[49]



- Arctic


The Arctic food web is complex. The loss of sea ice can ultimately affect the entire food web, from algae and plankton to fish to mammals. The impact of climate change on a particular species can ripple through a food web and affect a wide range of other organisms... Not only is the decline of sea ice impairing polar bear populations by reducing the extent of their primary habitat, it is also negatively impacting them via food web effects. Declines in the duration and extent of sea ice in the Arctic leads to declines in the abundance of ice algae, which thrive in nutrient-rich pockets in the ice. These algae are eaten by zooplankton, which are in turn eaten by Arctic cod, an important food source for many marine mammals, including seals. Seals are eaten by polar bears. Hence, declines in ice algae can contribute to declines in polar bear populations.[50]




Gray arrows: flow of carbon to heterotrophs
Green arrows: major pathways of carbon flow to or from mixotrophs
HCIL: heterotrophic ciliates; MCIL: mixotrophic ciliates; HNF: heterotrophic nanoflagellates; DOC: dissolved organic carbon; HDIN: heterotrophic dinoflagellates [51]
- Antarctic
-
 Antarctic jellyfish Diplulmaris antarctica under the ice
Antarctic jellyfish Diplulmaris antarctica under the ice -
![Colonies of the alga Phaeocystis antarctica, an important phytoplankter of the Ross Sea that dominates early season blooms after the sea ice retreats and exports significant carbon.[52]](https://upload.wikimedia.org/wikipedia/commons/thumb/a/a3/Phaeocystis.png/326px-Phaeocystis.png) Colonies of the alga Phaeocystis antarctica, an important phytoplankter of the Ross Sea that dominates early season blooms after the sea ice retreats and exports significant carbon.[52]
Colonies of the alga Phaeocystis antarctica, an important phytoplankter of the Ross Sea that dominates early season blooms after the sea ice retreats and exports significant carbon.[52] -
![The pennate diatom Fragilariopsis kerguelensis, found throughout the Antarctic Circumpolar Current, is a key driver of the global silicate pump.[53]](https://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Fragilariopsis_kerguelensis.jpg/248px-Fragilariopsis_kerguelensis.jpg) The pennate diatom Fragilariopsis kerguelensis, found throughout the Antarctic Circumpolar Current, is a key driver of the global silicate pump.[53]
The pennate diatom Fragilariopsis kerguelensis, found throughout the Antarctic Circumpolar Current, is a key driver of the global silicate pump.[53]

![Colonies of the alga Phaeocystis antarctica, an important phytoplankter of the Ross Sea that dominates early season blooms after the sea ice retreats and exports significant carbon.[52]](https://thcscience.wiki/term/cannabaceae/cannabis/cannabis-indica/?rdp_we_resource=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FFile%3APhaeocystis.png)
![The pennate diatom Fragilariopsis kerguelensis, found throughout the Antarctic Circumpolar Current, is a key driver of the global silicate pump.[53]](https://thcscience.wiki/term/cannabaceae/cannabis/cannabis-indica/?rdp_we_resource=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FFile%3AFragilariopsis_kerguelensis.jpg)

and carbon masses (Gt C) in dark boxes


Terrestrial comparisons

Compared to terrestrial biomass pyramids, aquatic pyramids are generally inverted at the base


Marine environments can have inversions in their biomass pyramids. In particular, the biomass of consumers (copepods, krill, shrimp, forage fish) is generally larger than the biomass of primary producers. This happens because the ocean's primary producers are mostly tiny phytoplankton which have r-strategist traits of growing and reproducing rapidly, so a small mass can have a fast rate of primary production. In contrast, many terrestrial primary producers, such as mature forests, have K-strategist traits of growing and reproducing slowly, so a much larger mass is needed to achieve the same rate of primary production.
Examples: The bristlecone pine can live for thousands of years, and has a very low production/biomass ratio. The cyanobacterium Prochlorococcus lives for about 24 hours, and has a very high production/biomass ratio.


In oceans, most primary production is performed by algae. This is a contrast to on land, where most primary production is performed by vascular plants.
Aquatic producers, such as planktonic algae or aquatic plants, lack the large accumulation of secondary growth that exists in the woody trees of terrestrial ecosystems. However, they are able to reproduce quickly enough to support a larger biomass of grazers. This inverts the pyramid. Primary consumers have longer lifespans and slower growth rates that accumulates more biomass than the producers they consume. Phytoplankton live just a few days, whereas the zooplankton eating the phytoplankton live for several weeks and the fish eating the zooplankton live for several consecutive years.[59] Aquatic predators also tend to have a lower death rate than the smaller consumers, which contributes to the inverted pyramidal pattern. Population structure, migration rates, and environmental refuge for prey are other possible causes for pyramids with biomass inverted. Energy pyramids, however, will always have an upright pyramid shape if all sources of food energy are included, since this is dictated by the second law of thermodynamics."[60][61]
Comparison of productivity in marine and terrestrial ecosystems [62]
| |||
|---|---|---|---|
| Ecosystem | Net primary productivity billion tonnes per year |
Total plant biomass billion tonnes |
Turnover time years |
Marine
|
45–55
|
1–2
|
0.02–0.06
|
Terrestrial
|
55–70
|
600–1000
|
9–20
|
Anthropogenic effects

- Overfishing
- Acidification
Pteropods and brittle stars together form the base of the Arctic food webs and both are seriously damaged by acidification. Pteropods shells dissolve with increasing acidification and brittle stars lose muscle mass when re-growing appendages.[64] Additionally the brittle star's eggs die within a few days when exposed to expected conditions resulting from Arctic acidification.[65] Acidification threatens to destroy Arctic food webs from the base up. Arctic waters are changing rapidly and are advanced in the process of becoming undersaturated with aragonite.[66] Arctic food webs are considered simple, meaning there are few steps in the food chain from small organisms to larger predators. For example, pteropods are "a key prey item of a number of higher predators – larger plankton, fish, seabirds, whales".[67]


- Climate change
"Our results show how future climate change can potentially weaken marine food webs through reduced energy flow to higher trophic levels and a shift towards a more detritus-based system, leading to food web simplification and altered producer–consumer dynamics, both of which have important implications for the structuring of benthic communities."[68][69]
"...increased temperatures reduce the vital flow of energy from the primary food producers at the bottom (e.g. algae), to intermediate consumers (herbivores), to predators at the top of marine food webs. Such disturbances in energy transfer can potentially lead to a decrease in food availability for top predators, which in turn, can lead to negative impacts for many marine species within these food webs... "Whilst climate change increased the productivity of plants, this was mainly due to an expansion of cyanobacteria (small blue-green algae)," said Mr Ullah. "This increased primary productivity does not support food webs, however, because these cyanobacteria are largely unpalatable and they are not consumed by herbivores. Understanding how ecosystems function under the effects of global warming is a challenge in ecological research. Most research on ocean warming involves simplified, short-term experiments based on only one or a few species."[69]


References
- ^ U S Department of Energy (2008) Carbon Cycling and Biosequestration page 81, Workshop report DOE/SC-108, U.S. Department of Energy Office of Science.
- ^ Campbell, Mike (22 June 2011). "The role of marine plankton in sequestration of carbon". EarthTimes. Retrieved 22 August 2014.
- ^ Why should we care about the ocean? NOAA: National Ocean Service. Updated: 7 January 2020. Retrieved 1 March 2020.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". In: Jungblut S., Liebich V., Bode-Dalby M. (Eds) YOUMARES 9-The Oceans: Our Research, Our Future , pages 279–297. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Karlson, A.M., Gorokhova, E., Gårdmark, A., Pekcan-Hekim, Z., Casini, M., Albertsson, J., Sundelin, B., Karlsson, O. and Bergström, L. (2020). "Linking consumer physiological status to food-web structure and prey food value in the Baltic Sea". Ambio, 49(2): 391–406. doi:10.1007/s13280-019-01201-1
- ^ Odum, W. E.; Heald, E. J. (1975) "The detritus-based food web of an estuarine mangrove community". Pages 265–286 in L. E. Cronin, ed. Estuarine research. Vol. 1. Academic Press, New York.
- ^ Pimm, S. L.; Lawton, J. H. (1978). "On feeding on more than one trophic level". Nature. 275 (5680): 542–544. doi:10.1038/275542a0.
- ^ Pauly, D.; Palomares, M. L. (2005). "Fishing down marine food webs: it is far more pervasive than we thought" (PDF). Bulletin of Marine Science. 76 (2): 197–211. Archived from the original (PDF) on 2013-05-14.
- ^ Cortés, E. (1999). "Standardized diet compositions and trophic levels of sharks". ICES J. Mar. Sci. 56 (5): 707–717. doi:10.1006/jmsc.1999.0489.
- ^ Pauly, D.; Trites, A.; Capuli, E.; Christensen, V. (1998). "Diet composition and trophic levels of marine mammals". ICES J. Mar. Sci. 55 (3): 467–481. doi:10.1006/jmsc.1997.0280.
- ^ a b Chlorophyll NASA Earth Observatory. Accessed 30 November 2019.
- ^ Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, Chen F, Lapidus A, Ferriera S, Johnson J, Steglich C, Church GM, Richardson P, Chisholm SW (December 2007). "Patterns and implications of gene gain and loss in the evolution of Prochlorococcus". PLoS Genetics. 3 (12): e231. doi:10.1371/journal.pgen.0030231. PMC 2151091. PMID 18159947.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Nemiroff, R.; Bonnell, J., eds. (27 September 2006). "Earth from Saturn". Astronomy Picture of the Day. NASA.
- ^ Partensky F, Hess WR, Vaulot D (March 1999). "Prochlorococcus, a marine photosynthetic prokaryote of global significance". Microbiology and Molecular Biology Reviews. 63 (1): 106–27. PMC 98958. PMID 10066832.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Mann, D. G. (1999). "The species concept in diatoms". Phycologia. 38: 437–495. doi:10.2216/i0031-8884-38-6-437.1.[permanent dead link]
- ^ Biology of Copepods Archived 2009-01-01 at the Wayback Machine at Carl von Ossietzky University of Oldenburg
- ^ a b c d Hays, G.C., Doyle, T.K. and Houghton, J.D. (2018) "A paradigm shift in the trophic importance of jellyfish?" Trends in ecology & evolution, 33(11): 874-884. doi:10.1016/j.tree.2018.09.001
- ^ a b Hamilton, G. (2016) "The secret lives of jellyfish: long regarded as minor players in ocean ecology, jellyfish are actually important parts of the marine food web". Nature, 531(7595): 432-435. doi:10.1038/531432a
- ^ Cardona, L., De Quevedo, I.Á., Borrell, A. and Aguilar, A. (2012) "Massive consumption of gelatinous plankton by Mediterranean apex predators". PloS one, 7(3): e31329. doi:10.1371/journal.pone.0031329
- ^ Tiny Forage Fish At Bottom Of Marine Food Web Get New Protections National Public Radio, 7 April 2016.
- ^ "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLoS ONE. 5 (10): e13255. 2010. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.
{{cite journal}}: Cite uses deprecated parameter|authors=(help)CS1 maint: unflagged free DOI (link) - ^ Brown, Joshua E. (12 Oct 2010). "Whale poop pumps up ocean health". Science Daily. Retrieved 18 August 2014.
- ^ Raina, J.B. (2018) "The life aquatic at the microscale". mSystems, 3(2): e00150-17. doi:10.1128/mSystems.00150-17.
- ^ Marco, D, ed. (2011). Metagenomics: Current Innovations and Future Trends. Caister Academic Press. ISBN 978-1-904455-87-5.
- ^ a b c Roslin, T. and Majaneva, S. (2016) "The use of DNA barcodes in food web construction—terrestrial and aquatic ecologists unite!". Genome, 59(9): 603–628. doi:10.1139/gen-2015-0229.
- ^ Wilhelm, Steven W.; Suttle, Curtis A. (1999). "Viruses and Nutrient Cycles in the Sea". BioScience. 49 (10): 781–788. doi:10.2307/1313569. ISSN 1525-3244. JSTOR 1313569.
- ^ Weinbauer, Markus G., et al. "Synergistic and antagonistic effects of viral lysis and protistan grazing on bacterial biomass, production and diversity." Environmental Microbiology 9.3 (2007): 777-788.
- ^ Robinson, Carol, and Nagappa Ramaiah. "Microbial heterotrophic metabolic rates constrain the microbial carbon pump." The American Association for the Advancement of Science, 2011.
- ^ Käse L, Geuer JK. (2018) "Phytoplankton responses to marine climate change–an introduction". In Jungblut S., Liebich V., Bode M. (Eds) YOUMARES 8–Oceans Across Boundaries: Learning from each other, pages 55–72, Springer. doi:10.1007/978-3-319-93284-2_5.
- ^ Heinrichs, M.E., Mori, C. and Dlugosch, L. (2020) "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". In: YOUMARES 9-The Oceans: Our Research, Our Future , pages 279–297. Springer. doi:10.1007/978-3-030-20389-4_15.
- ^ Gutierrez MH, Jara AM, Pantoja S (2016) "Fungal parasites infect marine diatoms in the upwelling ecosystem of the Humboldt current system off central Chile". Environ Microbiol, 18(5): 1646–1653. doi:10.1111/1462-2920.13257.
- ^ Coll, M., Schmidt, A., Romanuk, T. and Lotze, H.K. (2011). "Food-web structure of seagrass communities across different spatial scales and human impacts". PloS ONE, 6(7): e22591. doi:10.1371/journal.pone.0022591.
- ^ Heymans, J.J., Coll, M., Libralato, S., Morissette, L. and Christensen, V. (2014). "Global patterns in ecological indicators of marine food webs: a modelling approach". PloS one, 9(4). doi:10.1371/journal.pone.0095845.
- ^ Pranovi, F., Libralato, S., Raicevich, S., Granzotto, A., Pastres, R. and Giovanardi, O. (2003). "Mechanical clam dredging in Venice lagoon: ecosystem effects evaluated with a trophic mass-balance model". Marine Biology, 143(2): 393–403. doi:10.1007/s00227-003-1072-1.
- ^ Leray M, Meyer CP, Mills SC. (2015) "Metabarcoding dietary analysis of coral dwelling predatory fish demonstrates the minor contribution of coral mutualists to their highly partitioned, generalist diet". PeerJ, 3: e1047. doi:10.7717/peerj.1047.
- ^ Wang, F., Wu, Y., Chen, Z., Zhang, G., Zhang, J., Zheng, S. and Kattner, G. (2019) "Trophic interactions of mesopelagic fishes in the South China Sea illustrated by stable isotopes and fatty acids". Frontiers in Marine Science, 5: 522. doi:10.3389/fmars.2018.00522.
- ^ a b Tollefson, Jeff (27 February 2020) Enter the twilight zone: scientists dive into the oceans’ mysterious middle Nature News. doi:10.1038/d41586-020-00520-8.
- ^ Irigoien, X., Klevjer, T.A., Røstad, A., Martinez, U., Boyra, G., Acuña, J.L., Bode, A., Echevarria, F., Gonzalez-Gordillo, J.I., Hernandez-Leon, S. and Agusti, S. (2014) "Large mesopelagic fishes biomass and trophic efficiency in the open ocean". Nature communications, 5: 3271. doi:10.1038/ncomms4271
- ^ Fish biomass in the ocean is 10 times higher than estimated EurekAlert, 7 February 2014.
- ^ Choy, C.A., Wabnitz, C.C., Weijerman, M., Woodworth-Jefcoats, P.A. and Polovina, J.J. (2016) "Finding the way to the top: how the composition of oceanic mid-trophic micronekton groups determines apex predator biomass in the central North Pacific". Marine Ecology Progress Series, 549: 9–25. doi:10.3354/meps11680.
- ^ Choy, C.A., Haddock, S.H. and Robison, B.H. (2017) "Deep pelagic food web structure as revealed by in situ feeding observations". Proceedings of the Royal Society B: Biological Sciences, 284(1868): 20172116. doi:10.1098/rspb.2017.2116.
- ^ Kelly, T.B., Davison, P.C., Goericke, R., Landry, M.R., Ohman, M. and Stukel, M.R. (2019) "The importance of mesozooplankton diel vertical migration for sustaining a mesopelagic food web". Frontiers in Marine Science, 6: 508. doi:10.3389/fmars.2019.00508.
- ^ Levin LA, Baco AR, Bowden DA, Colaco A, Cordes EE, Cunha MR, Demopoulos AWJ, Gobin J, Grupe BM, Le J, Metaxas A, Netburn AN, Rouse GW, Thurber AR, Tunnicliffe V, Van Dover CL, Vanreusel A and Watling L (2016). "Hydrothermal Vents and Methane Seeps: Rethinking the Sphere of Influence". Front. Mar. Sci. 3:72. doi:10.3389/fmars.2016.00072
- ^ Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L. and Sarrazin, J. (2016). "Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps". PloS ONE, 11(9): p.e0162263. doi:10.1371/journal.pone.0162263.
- ^ Bernardino AF, Levin LA, Thurber AR and Smith CR (2012). "Comparative Composition, Diversity and Trophic Ecology of Sediment Macrofauna at Vents, Seeps and Organic Falls". PLoS ONE, 7(4): e33515. pmid:22496753. doi:10.1371/journal.pone.0033515.
- ^ Portail M, Olu K, Escobar-Briones E, Caprais JC, Menot L, Waeles M, et al. (2015). "Comparative study of vent and seep macrofaunal communities in the Guaymas Basin". Biogeosciences. 12(18): 5455–79. doi:10.5194/bg-12-5455-2015.
- ^ McCarthy, J.J., Canziani, O.F., Leary, N.A., Dokken, D.J. and White, K.S. (Eds.) (2001) Climate Change 2001: Impacts, Adaptation, and Vulnerability: Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change Page 807, Cambridge University Press. ISBN 9780521015004
- ^ Murphy, E.J., Cavanagh, R.D., Drinkwater, K.F., Grant, S.M., Heymans, J.J., Hofmann, E.E., Hunt Jr, G.L. and Johnston, N.M. (2016) "Understanding the structure and functioning of polar pelagic ecosystems to predict the impacts of change". Proceedings of the Royal Society B: Biological Sciences, 283(1844): 20161646. doi:10.1098/rspb.2016.1646.
- ^ Climate Impacts on Ecosystems: Food Web Disruptions EPA. Accessed 11 February 2020.
- ^ Stoecker, D.K. and Lavrentyev, P.J. (2018). "Mixotrophic plankton in the polar seas: a pan-arctic review". Frontiers in Marine Science, 5: 292. doi:10.3389/fmars.2018.00292
- ^ Bender, S.J., Moran, D.M., McIlvin, M.R., Zheng, H., McCrow, J.P., Badger, J., DiTullio, G.R., Allen, A.E. and Saito, M.A. (2018) "Colony formation in Phaeocystis antarctica: connecting molecular mechanisms with iron biogeochemistry". Biogeosciences, 15(16): 4923–4942. doi:10.5194/bg-15-4923-2018.
- ^ Pinkernell, S. and Beszteri, B. (2014) "Potential effects of climate change on the distribution range of the main silicate sinker of the Southern Ocean". Ecology and evolution, 4(16): 3147–3161. doi:10.1002/ece3.1138
- ^ Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". Nature communications, 10(1): 1–13. doi:10.1038/s41467-019-12668-7.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b Cordone, G., Marina, T.I., Salinas, V., Doyle, S.R., Saravia, L.A. and Momo, F.R.(2018). "Effects of macroalgae loss in an Antarctic marine food web: applying extinction thresholds to food web studies". PeerJ, 6: e5531. doi:10.7717/peerj.5531
- ^ Marina, T.I., Salinas, V., Cordone, G., Campana, G., Moreira, E., Deregibus, D., Torre, L., Sahade, R., Tatian, M., Oro, E.B. and De Troch, M. (2018). "The food web of Potter Cove (Antarctica): complexity, structure and function". Estuarine, Coastal and Shelf Science, 200: 141–151. doi:10.1016/j.ecss.2017.10.015.
- ^ "Oldlist". Rocky Mountain Tree Ring Research. Retrieved January 8, 2013.
- ^ Bar-On, Y.M., Phillips, R. and Milo, R. (2018) "The biomass distribution on Earth". Proceedings of the National Academy of Sciences, 115(25): 6506–6511. doi:10.1073/pnas.1711842115.
- ^ Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. p. 167. ISBN 978-1-4200-5544-3.
- ^ Odum, E. P.; Barrett, G. W. (2005). Fundamentals of Ecology (5th ed.). Brooks/Cole, a part of Cengage Learning. ISBN 978-0-534-42066-6. Archived from the original on 2011-08-20.
- ^ Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). "Modeling inverted biomass pyramids and refuges in ecosystems" (PDF). Ecological Modelling. 220 (11): 1376–1382. doi:10.1016/j.ecolmodel.2009.03.005. Archived from the original (PDF) on 2011-10-07.
- ^ Field, C.B., Behrenfeld, M.J., Randerson, J.T. and Falkowski, P. (1998) "Primary production of the biosphere: integrating terrestrial and oceanic components". Science, 281(5374): 237–240. doi:10.1126/science.281.5374.237.
- ^ Maureaud, A., Gascuel, D., Colléter, M., Palomares, M.L., Du Pontavice, H., Pauly, D. and Cheung, W.W. (2017) "Global change in the trophic functioning of marine food webs". PloS one, 12(8): e0182826. doi:10.1371/journal.pone.0182826
- ^ "Effects of Ocean Acidification on Marine Species & Ecosystems". Report. OCEANA. Retrieved 13 October 2013.
- ^ "Comprehensive study of Arctic Ocean acidification". Study. CICERO. Archived from the original on 10 December 2013. Retrieved 14 November 2013.
- ^ Lischka, S.; Büdenbender J.; Boxhammer T.; Riebesell U. (15 April 2011). "Impact of ocean acidification and elevated temperatures on early juveniles of the polar shelled pteropod Limacina helicina : mortality, shell degradation, and shell growth" (PDF). Report. Biogeosciences. pp. 919–932. Retrieved 14 November 2013.
- ^ "Antarctic marine wildlife is under threat, study finds". BBC Nature. Retrieved 13 October 2013.
- ^ Ullah, H., Nagelkerken, I., Goldenberg, S.U. and Fordham, D.A. (2018) "Climate change could drive marine food web collapse through altered trophic flows and cyanobacterial proliferation". PLoS biology, 16(1): e2003446. doi:10.1371/journal.pbio.2003446
- ^ a b Climate change drives collapse in marine food webs ScienceDaily. 9 January 2018.
- ^ Luypaert, T., Hagan, J.G., McCarthy, M.L. and Poti, M. (2020) "Status of Marine Biodiversity in the Anthropocene". In: YOUMARES 9-The Oceans: Our Research, Our Future, pages 57-82, Springer. doi:10.1007/978-3-030-20389-4_4.
- ^ IUCN (2018) The IUCN Red List of Threatened Species: Version 2018-1
- ^ Luypaert, T., Hagan, J.G., McCarthy, M.L. and Poti, M. (2020) "Status of Marine Biodiversity in the Anthropocene". In: YOUMARES 9-The Oceans: Our Research, Our Future, pages 57-82, Springer. doi:10.1007/978-3-030-20389-4_4.
- ^ IUCN (2018) The IUCN Red List of Threatened Species: Version 2018-1